Spinal Cord Injury
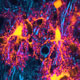
 surrounded by a wall of reactive astrocytes (white).")
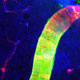
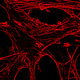
Axial cross section of rodent spinal cord following acute contusion injury. Depicted in the center is the glial scar, consisting of an inner core of fibroblasts (blue) surrounded by a wall of reactive astrocytes (white).
Traumatic spinal cord injury (SCI) is a devastating assault to the central nervous system (CNS) that often results in permanent neurologic impairment, intense personal suffering and a disruption to essentially every aspect of life. An overwhelming majority of patients that suffer spinal cord injuries also develop chronic neuropathic pain syndromes that often persist indefinitely. Yet, despite high prevalence and decades of intensive study, to date, damage sustained from spinal cord injury is largely irreversible.
Pathologically, SCI manifests in two phases: the acute events surrounding primary injury typically result in damage or disturbance to the neuronal elements of the spinal cord, leading to the severing of axon tracts and neuronal cell death; however, within 1-3 days of the initial injury, SCI patients often experience a cascade of secondary pathological changes that amplify the initial traumatic injury and result in edema, spreading necrosis, inflammation, ischemia, and proliferation of reactive glial cells. Although little is known about the pathogenesis of this secondary phase, histological study has revealed a distinctive pattern of cellular events: in what appears to be a defense mechanism aimed at halting the spread of cellular damage, the injured neural tissue becomes isolated from surrounding healthy tissue by the formation of what is commonly referred to as the glial scar
, consisting of a dense core of fibroblasts, surrounded by a symmetrical wall of reactive astrocytes.

Spinal cord injury induced Glial scar.
Following SCI, astrocytes – the largest and most abundant glial cell in the CNS – undergo reactive changes and form a barrier of projections linked together by gap-junctions. The hallmark of reactive changes in astrocytes, also known as “astrogliosis”, is the upregulation of the expression of intermediate filament proteins (e.g. glial-fibrillary-acid protein), and gap junction proteins (e.g. connexin 43) that together facilitate network communication and the passage of small intracellular molecules from one astrocyte to the next.
Research in our lab has shown that astrocytes also engage in long distance signaling via the release of purines, such as adenosine tri-phosphate (ATP), which non-selectively stimulates purinergic receptors on a number of cell types, including neurons and microglial cells. Although the cellular source of ATP remains a matter of some controversy, bioluminescent imaging of ATP in animal models suggests that large quantities are released to the extracellular space following acute spinal cord injury. Our work has focused on the role that purinergic signaling may play in the pathogenesis of the early secondary expansion of injury and the onset of conditions that prevent the regrowth of injured axons, as this time period represents a unique opportunity to reduce the severity of tissue injury. Stimulation of one purinergic (ATP) receptor in particular, P2X7, on microglia has been shown to trigger the release of a host of additional signaling molecules, including proinflammatory cytokines and agents linked to the development of chronic neuropathic pain.
Understanding the mechanisms of the secondary phase of spreading cellular damage following acute SCI constitutes an important step that will hopefully translate to developing treatments with specific cellular and molecular targets. In this regard, we have developed a small P2X7 receptor antagonist derived from harmless blue food coloring (Brilliant blue G) that has been shown to block the local action of ATP and disrupt the sequelae of its downstream signaling consequences. Ongoing work focuses on identifying additional pharmacological agents suitable for translation to the clinical setting.
This project is supported by funding from DOD – Spinal Cord Injury Research Program (Development of a Small Molecule P2X7R Antagonist as a Treatment for Acute SCI
) & NIH – National Institutes of Neurological Disorders and Stroke. (ATP as the Instigator of Inflammatory Responses to Spinal Cord Injury
).
Further Reading
Critical role of connexin 43 in secondary expansion of traumatic spinal cord injury. Huang C, Han X, Li X, Lam E, Peng W, Lou N, Torres A, Yang M, Garre JM, Tian GF, Bennett MV, Nedergaard M, Takano T. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2012 Mar 7; 32(10):3333-8.
Astrocytic CX43 hemichannels and gap junctions play a crucial role in development of chronic neuropathic pain following spinal cord injury. Chen MJ, Kress B, Han X, Moll K, Peng W, Ji RR, Nedergaard M. Glia. 2012 Nov 0; 60(11):1660-70. Epub 2012 Aug 01.
Connexin and pannexin hemichannels in inflammatory responses of glia and neurons. Bennett MV, Garré JM, Orellana JA, Bukauskas FF, Nedergaard M, Sáez JC. Brain research. 2012 1487:3-15. Epub 2012 Sep 10.
Functions of astrocytes and their potential as therapeutic targets. Kimelberg HK, Nedergaard M. Neurotherapeutics : the journal of the American Society for Experimental NeuroTherapeutics. 2010 Oct 0; 7(4):338-53.
Physiological and pathological functions of P2X7 receptor in the spinal cord. Cotrina ML, Nedergaard M. Purinergic signalling. 2009 Jun 0; 5(2):223-32. Epub 2009 Feb 11.
Systemic administration of an antagonist of the ATP-sensitive receptor P2X7 improves recovery after spinal cord injury. Peng W, Cotrina ML, Han X, Yu H, Bekar L, Blum L, Takano T, Tian GF, Goldman SA, Nedergaard M. Proceedings of the National Academy of Sciences of the United States of America. 2009 Jul 28; 106(30):12489-93. Epub 2009 Jul 27.
Connexin 43 hemichannels are permeable to ATP. Kang J, Kang N, Lovatt D, Torres A, Zhao Z, Lin J, Nedergaard M. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2008 Apr 30; 28(18):4702-11.
P2X7 receptor inhibition improves recovery after spinal cord injury. Wang X, Arcuino G, Takano T, Lin J, Peng WG, Wan P, Li P, Xu Q, Liu QS, Goldman SA, Nedergaard M. Nature medicine. 2004 Aug 0; 10(8):821-7. Epub 2004 Jul 18.